






L’origine de la zoonose et les modes de contamination
Publié en ligne le 13 avril 2020 - Covid-19 -
1 | Brugère-Picoux J, « Covid-19 : origine de la zoonose et modes de contamination », 17 mars 2020, sur www.fondation-droit-animal.org
La nouvelle maladie émergente Covid-19 (Coronavirus disease 2019) est due à un coronavirus (Sars-CoV-2) dont la chauve-souris est un réservoir naturel. Non affecté, ce mammifère volant peut transmettre de nombreux virus à d’autres espèces, dont l’espèce humaine.
Les chauves-souris, réservoirs de virus émergents
Les chiroptères, plus communément appelés chauves-souris, seuls mammifères volants, constituent environ 20 % de la diversité des mammifères et sont largement distribués dans le monde. Ils présentent une longévité de plusieurs dizaines d’années, inhabituelle pour des animaux de petite taille au métabolisme élevé [1]. Depuis quelques décennies, on découvre que ces chauves-souris peuvent héberger de nombreux virus dont certains sont à l’origine de maladies émergentes chez l’Homme ou l’animal. Ces réservoirs naturels de virus présentent aussi la particularité de ne pas être affectés par les agents infectieux qu’ils hébergent [2]. On a pu montrer que les chauves-souris étaient résistantes à des charges virales, mortelles pour d’autres mammifères, d’Henipavirus et de Lyssavirus [3,4]. Cependant on connaît mal les mécanismes responsables de cette capacité des chauves-souris à coexister avec les virus [5]. Leur adaptation au vol favoriserait une élévation de leur température corporelle permettant d’accroître l’efficacité de la réponse immunitaire [6,7]. Cette coexistence entre virus et chauves-souris serait aussi liée à une réponse atténuée en interférons [8] qui permettrait le maintien d’une certaine charge virale en permanence dans l’organisme.
Lors de l’émergence d’une maladie virale impliquant des chauves-souris, le franchissement occasionnel de la barrière d’espèce de la chauve-souris à l’Homme ou à l’animal est souvent lié à une modification des écosystèmes largement due à des activités humaines. Alors qu’il n’existait que peu de contacts auparavant, les nouvelles conditions permettent la mise en relation des espèces animales sauvages (chauve-souris ou hôtes intermédiaires) avec l’Homme ou certaines espèces animales domestiques.
C’est ainsi que les chauves-souris ont été les pourvoyeuses de maladies zoonotiques émergentes redoutables déclenchées par divers agents viraux dont les Lyssavirus, les Henipavirus, les Filovirus et certains coronavirus (voir encadré).
Les chauves-souris ont été les pourvoyeuses de maladies zoonotiques émergentes redoutables déclenchées par divers agents viraux dont :
- les Lyssavirus, groupe auquel appartient le virus de la rage : les réservoirs animaux sont actuellement les chauves-souris et les carnivores, les chauves-souris constituant probablement le réservoir originel [1] ;
- les Henipavirus, parmi lesquels : le virus Hendra rencontré en Australie depuis 1994 [2], responsable d’une affection heureusement rare car souvent mortelle chez l’Homme et le cheval ; le virus Nipah, identifié en 1998 lors d’une épidémie affectant des fermes porcines en Malaisie dont les porcs présentaient des troubles respiratoires mais aussi les personnes en contact avec les porcs (265 malades dont 105 décès à la suite d’une encéphalite) [3]. C’est un peu plus tard que l’on a découvert que des chauves-souris frugivores (Pteropus sp.) étaient à l’origine de cette épidémie qui s’est étendue au Bangladesh et en Inde, notamment par la consommation du jus de palme contaminé [4] ;
- les Filovirus, parmi lesquels : le virus de Marburg décrit tout d’abord en Allemagne en 1967 à la suite de contaminations de laborantins travaillant sur des cellules rénales de singes verts [5], puis découvert chez les chauves-souris [6-8] suite à des contaminations humaines mortelles consécutives à la visite en 1980 de grottes qui en hébergeaient [9] ; le virus Ebola pour lequel le rôle réservoir de la chauve-souris fut démontré en 2005 [10-12] ;
- certains coronavirus et plus spécifiquement des Betacoronavirus, comme le virus du syndrome respiratoire aigu sévère (Sras), celui du syndrome respiratoire du Moyen-Orient (Mers), ou celui du Coronavirus disease 2019 (Covid-19).
1 | Bourhy H, « De la négligence à la réémergence de la rage », in Brugère-Picoux J, Rey M, Les maladies infectieuses exotiques. Risques d’importation et d’implantation en Europe, Rapports de l’Académie nationale de Médecine. Éditions Lavoisier, 2010, p. 117-32.
2 | Selvey LA et al., “Infection of humans and horses by a newly described morbillivirus”, Medical Journal of Australia,1995,162 : 642-5.
3 | Chua KB et al., “Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia”, Lancet, 1999, 354:1257-9.
4 | Luby SP et al., “Foodborne transmission of Nipah virus, Bangladesh”, Emerg. Infect. Dis., 2006, 12:1888-94.
5 | Smith CE et al., “Fatal human disease from vervet monkeys”, Lancet, 1967, 2:1119-21.
6 | Towner JS et al., “Marburg Virus Infection Detected in a Common African Bat”, PLoS ONE, 2007, 2 : e764.
7 | Towner JS et al., “Isolation of Genetically Diverse Marburg Viruses from Egyptian Fruit Bats”, PLoS Pathos., 2009, 5:e1000536.
8 | Maganga GD et al., “Is Marburg Virus Enzootic in Gabon ?”, J. Inf. Dis., 2011 ;204:S800-3.
9 | Smith DH et al., “Marburg-virus disease in Kenya”, Lancet, 1982, 1:816-20.
10 | Leroy EM et al., “Fruit bats as reservoirs of Ebola virus”, Nature, 2005, 438:575-6.
11 | Leroy EM et al., “Human Ebola Outbreak Resulting from Direct Exposure to Fruit Bats in Luebo, Democratic Republic of Congo”, Vector-Borne and Zoonotic Diseases, 2009, 9:723-8.
12 | Pourrut X et al., “Large serological survey showing cocirculation of Ebola and Marburg viruses in Gabonese bat populations, and a high seroprevalence of both viruses in Rousettus aegyptiacus”, BMC Infectious Diseases, 2009, 9:159.
L’origine des coronavirus
Les coronavirus sont des virus à ARN classés en Alphacoronavirus, Betacoronavirus, Gammacoronavirus et Deltacoronavirus. Leur nom vient de l’observation de petites excroissances (ou spicules) à leur surface, formant une sorte de couronne.
Les coronavirus semblent provenir des chauves-souris, et plus particulièrement les Alphacoronavirus et les Betacoronavirus (genres pour lesquels on constate des zoonoses), ce qui explique le grand nombre de virus isolés dans ces espèces alors que les oiseaux seraient à l’origine des Gammacoronavirus et des Deltacoronavirus (cf. figure 1) [9].
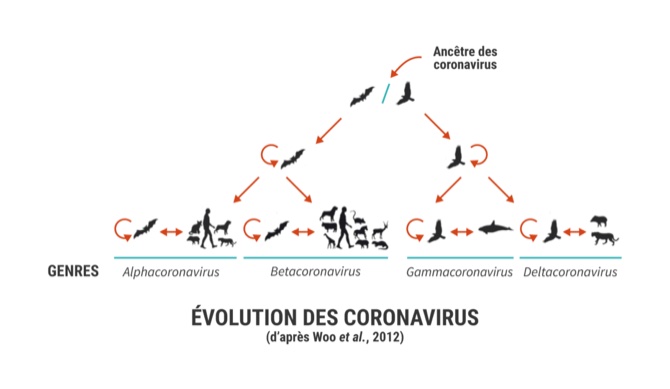
Le premier coronavirus identifié fut celui de la bronchite infectieuse aviaire en 1931 aux États-Unis alors que ce n’est que dans les années 1960 que les premiers coronavirus ont été décrits chez l’Homme, responsables de rhumes souvent banals. Depuis, de nombreux coronavirus ont été isolés chez les Mammifères et les Oiseaux.
La plupart des Alphacoronavirus sont spécifiques d’espèces. Ils peuvent être responsables de maladies graves comme la péritonite infectieuse féline chez le chat ou la gastroentérite transmissible (GET) chez le porcelet. L’émergence en 1984 du coronavirus respiratoire porcin semble avoir pour origine une modification du virus de la GET [10]. La GET est devenue plus rare, parallèlement à l’émergence d’un autre coronavirus (Swine acute diarrhea syndrome coronavirus ou Sads-CoV), responsable de la diarrhée épidémique porcine (DEP) surtout importante dans le Sud-Est asiatique depuis 2010 [11]. En 2017, une importante épidémie chez les porcelets a permis de démontrer que le réservoir animal de cette diarrhée épidémique porcine était vraisemblablement la chauve-souris [12].
Le Sars-CoV-1 est un Betacoronavirus responsable du syndrome respiratoire aigu sévère (Sras) qui fut une importante épidémie de février à mai 2003, avec un taux de mortalité de 10 %, tuant 774 personnes sur 8096 malades, surtout en Chine mais le Canada fut aussi très touché (avec 43 décès sur 251 malades) [13]. Il a fallu mettre en place d’importantes mesures de biosécurité pour assister à la fin de l’épidémie. Quand le Sras a émergé à la mi-novembre 2002 dans la province du Guangdong, les cas n’ont pas été officiellement notifiés par crainte d’éventuelles retombées sociales ou économiques, permettant ainsi une large diffusion du virus. L’Organisation mondiale de la santé (OMS) n’a été prévenue que le 11 février 2003…
Le Mers est apparu plus tard en septembre 2012 et concerne principalement le Moyen-Orient, l’animal réservoir étant le dromadaire. À la fin de novembre 2019, 2 494 cas ont été confirmés dont 858 sont décédés (soit un taux de mortalité de 34,4 %) [14]. L’Arabie Saoudite a été le pays le plus touché avec 2102 cas dont 780 sont décédés, soit un taux de mortalité de 37,1 %.
Alors que les premiers cas de Sras ont été observés en 2002 dans la province du Guangdong, il s’avère que la source géographique du virus semble être la province de Yunnan ou le sud-ouest de la Chine, le principal réservoir animal étant vraisemblablement des chauves-souris fer à cheval (Rhinolophus sinicus) [2],[15],[16]. Une surveillance de ces chauves-souris réalisée sur plus de cinq années dans une grotte de la province de Yunnan a permis de démontrer l’importante quantité de coronavirus pouvant être hébergés par ces chiroptères dont certains proches du virus du Sras (Sars-CoV-1) et dénommés Sars-related coronavirus (Sarsr-CoV). Les scientifiques chinois soulignèrent même en 2017 que ces nouvelles informations sur l’origine et l’évolution du Sars-CoV-1 mettaient en évidence la nécessité de se préparer à l’émergence future de maladies comme le Sras [17]…
D’autres scientifiques américains [18] et chinois [19] avaient signalé (en 2015 et 2019 respectivement) le potentiel d’émergence des coronavirus présents dans les populations de chauves-souris, en particulier le coronavirus Shco14-CoV circulant couramment chez les chauves-souris fer à cheval du fait de sa réplication identique au Sars-CoV-1 dans des cellules primaires de poumon humain. Ainsi, les scientifiques chinois concluaient : « Il est très probable que les futures épidémies de coronaviroses comme le Sras ou le Mers proviendront de chauves-souris, et il y a une probabilité accrue que cela se produise en Chine. Par conséquent, l’enquête sur les coronavirus de chauve-souris devient un problème urgent pour la détection des signes d’alerte précoce pour minimiser alors l’impact de ces futures épidémies en Chine »… Il était surtout évident pour ces scientifiques que la Chine représentait la zone à haut risque (le point chaud) d’où partirait l’épidémie.
Ces alertes scientifiques américaines et chinoises de 2015 et de 2019 n’ont pas permis d’éviter l’apparition en décembre 2019 d’une pneumonie d’origine inconnue touchant 59 personnes dans la ville chinoise de Wuhan. Les personnes atteintes avaient surtout fréquenté le marché de la ville où l’on vendait divers animaux domestiques et sauvages le plus souvent vivants. Le 2 janvier 2020, ce marché fut immédiatement fermé sans que l’on ait recherché l’origine de la contamination parmi les espèces animales vendues. Cette maladie émergente (dénommée Coronavirus disease 2019 ou Covid-19) est due à un coronavirus (Sars-CoV-2) où une autre chauve-souris fer à cheval (Rhinolophus affinis) est de nouveau incriminée en tant que réservoir. L’étude du génome du Sars-CoV-2 [20] confirme qu’il s’agit d’un virus proche à 96,2 % d’un coronavirus présent chez la chauve-souris (Sars-CoV ; RaTG13), ce virus étant plus éloigné du virus du Sras (79 %) ou de celui du Mers (50 %).
L’origine de la contamination par les coronavirus
On ne connaît pas l’origine exacte de la contamination humaine par le Sars-CoV-2 alors que l’on connaît le lien épidémiologique avec un marché aux animaux vivants sauvages ou domestiques qui n’est pas sans rappeler l’origine du Sras également liée à un marché d’animaux vivants en Chine. La question n’est actuellement pas résolue et l’on ne peut que se baser sur les études réalisées sur le Sars-CoV-1. On peut regretter que des prélèvements n’aient pas été réalisés sur les animaux (en particulier les animaux sauvages) vendus vivants dans une grande proximité sur le marché de Wuhan, malgré l’illégalité de leur vente, avant la fermeture de ce dernier.
On peut retenir trois hypothèses à l’origine du franchissement d’un coronavirus de la barrière d’espèce de la chauve-souris vers l’Homme (figure 2).
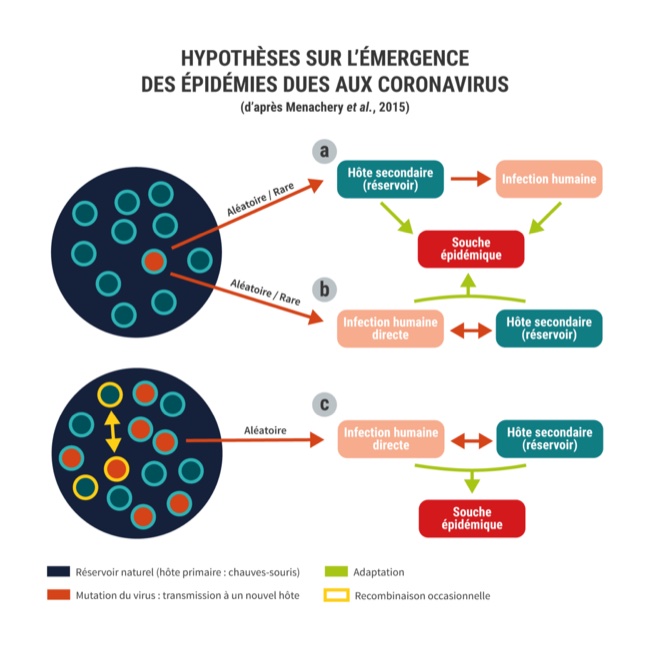
Comme tous les virus à ARN de manière générale, les coronavirus sont connus pour avoir un taux de mutation élevé pendant la réplication et sont sujets à la recombinaison si différents virus infectent le même individu. Les populations de chauves-souris hébergent plusieurs types de coronavirus (cercle bleu foncé à contour bleu clair). Le Sars-CoV serait la conséquence d’une mutation au sein de l’hôte (cercle orange) ayant permis de façon aléatoire et rare l’émergence d’une souche chez un nouvel hôte.
La première hypothèse (a) concerne un hôte intermédiaire comme la civette (souche civette) ayant permis ultérieurement la contamination humaine. Cependant les premières souches humaines du Sars-CoV-1 étant plus étroitement liées aux souches chauves-souris qu’aux souches civettes, il est possible que la contamination humaine ait été directe à partir de la chauve-souris (hypothèse b), les civettes masquées ayant joué un rôle réservoir secondaire favorisant le maintien de l’infection animale et humaine dans les marchés d’animaux vivants. Une troisième hypothèse (c) est la présence de multiples virus proches du Sras n’ayant pas subi de mutation et hébergés par les chauves-souris. Une adaptation secondaire ou une recombinaison avec une protéine de pointe lui permettant de se lier aux récepteurs cellulaires humains (cercle de contour jaune) peut favoriser une contamination directe de l’Homme.
[1] Menachery VD et al., “A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence”, Nature Medicine, 2015, 21:1508-13.
Virus mutant avec nécessité d’un hôte intermédiaire pour infecter l’Homme
Si l’on se rappelle l’épidémie du Sras, des civettes palmistes masquées (Paguma larvata) d’origine sauvage ou provenant de fermes d’élevage, vendues vivantes sur les marchés, furent rapidement incriminées en tant que réservoirs de l’agent pathogène et il s’ensuivit un abattage massif de ces petits Viverridae par mesure de précaution avant que l’on ne découvre que la chauve-souris était le réservoir principal du Sars-CoV-1. A alors émergé l’hypothèse selon laquelle la civette, contaminée par la chauve-souris, avait été l’hôte intermédiaire qui a ultérieurement contaminé l’Homme.
D’autres espèces présentes sur les marchés d’animaux vivants se sont révélées porteuses du Sars-CoV-1 sans être retenues en tant qu’hôtes intermédiaires à l’origine du Sras [21] : le chien viverrin (Nyctereutes procyonoides) surtout recherché pour sa fourrure et le blaireau furet (Melogale moschata) [22,23], ainsi que les renards roux, les chats domestiques et les rats [24], ces espèces ayant pu avoir été contaminées par les civettes palmistes masquées, voire l’Homme sur les marchés (ou inversement).
Rappelons que d’autres animaux ont pu être infectés expérimentalement par le Sars-CoV-1 : singes, chats et furets [25], souris[26-29], cobayes [29], hamsters [30].

Dans le cas du Covid-19, le seul animal actuellement suspecté en tant qu’hôte intermédiaire potentiel entre la chauve-souris et l’Homme est le pangolin (Manis pentadactyla et Manis javanica en Asie). Le pangolin, de l’ordre des Pholidotes, vit dans les forêts en se nourrissant de termites et de fourmis. Cet animal sauvage est apprécié en Asie pour sa viande mais aussi pour ses écailles qui auraient des propriétés thérapeutiques en médecine traditionnelle asiatique. Il est très braconné dans le monde alors que la Convention internationale sur le commerce d’espèces sauvages menacées d’extinction (Cites) a voté en 2017 l’interdiction totale du commerce international des pangolins (voir sur le site de l’International Union for Conservation of Nature (IUCN) [31]). L’avantage de la suspicion concernant le pangolin dans le Covid-19 sera un contrôle plus strict de son commerce permettant de le protéger.
La possibilité d’un éventuel portage du Sars-CoV-2 par les animaux de compagnie a été évoquée lorsque l’on a découvert à Hong Kong fin février un chien positif dont la propriétaire était malade. Le suivi de ce chien a permis de noter un très faible taux de virus dans les voies respiratoires supérieures et l’absence d’anticorps sériques (un autre examen sera réalisé après quatorze jours d’observation) permettant de penser que l’infection a été modérée et que la contamination s’est effectuée de la propriétaire vers le chien [32]. Jusqu’à présent, il n’a jamais été observé que le Sars-CoV-1 pouvait être transmis d’un animal de compagnie (chien ou chat) vers l’Homme.
Virus mutant infectant directement l’Homme
Le rôle d’hôte intermédiaire de la civette palmiste masquée reste cependant hypothétique : si plusieurs cas de contamination humaine semblent avérés à partir de civettes palmistes masquées [23,33,34] dont certains cas bénins de Sras réapparus en 2004 [22,23,28,35], il s’avère que des civettes palmistes masquées d’origine sauvage pouvaient être négatives [36] comme d’autres civettes dans des fermes d’élevage [23,37]. Certaines civettes ont pu être contaminées dans le marché d’animaux vivants [23] ou dans les élevages souvent en surdensité animale avec un mélange de diverses espèces animales [37].
Il s’agirait alors de la deuxième hypothèse, à savoir une contamination directe de l’Homme à partir de la chauve-souris, les civettes masquées ayant joué un rôle réservoir secondaire favorisant le maintien de l’infection animale et humaine dans les marchés d’animaux vivants. Cette hypothèse est plausible car une analyse phylogénétique [38] a montré que les premières souches humaines du Sars-CoV-1 étaient plus étroitement liées aux souches chauves-souris qu’aux souches civettes.
En Chine, les animaux sauvages et domestiques sont souvent vendus vivants sur les marchés (ils sont parfois abattus au moment de la vente). La promiscuité de ces différentes espèces peut favoriser des transferts inter-espèces et parfois une contamination de l’Homme. Cela a été démontré pour une autre maladie virale rencontrée en Chine : les cas humains dus au virus aviaire influenza H7N9 observés ont pu diminuer drastiquement en 2013 avec l’interdiction des marchés de volailles vivantes à l’origine des contaminations [39].
Adaptation ou recombinaison du virus permettant d’infecter directement l’Homme
La troisième hypothèse permet de suspecter une contamination directe de l’Homme à partir de la chauve-souris à la suite d’une recombinaison d’un Sarsr-CoV présent chez l’animal. La recombinaison pourrait avoir eu lieu au niveau de la protéine S (dénommée « spike » du fait de sa forme en pointe).
En Chine, la plupart des chauves-souris hébergeant les Sarsr-CoV se retrouvent surtout dans certaines zones rurales du sud de la Chine (province de Yunnan) où les habitations sont proches de grottes abritant des Chiroptères. Une étude sérologique montrant 2,7 % de séropositivité chez 218 personnes vivant à proximité de ces grottes permet de suggérer la possibilité d’une contamination directe [40].
Les chauves-souris, consommées régulièrement, peuvent être vendues vivantes sur des marchés, favorisant ainsi les franchissements des barrières d’espèce, notamment vers l’Homme. Le risque est cependant plus élevé pour le chasseur, manipulant les animaux avec le risque de morsure ou de contact avec leur sang, que pour l’acheteur. Les contacts fréquents entre l’Homme et des chauves-souris vivantes peuvent être aussi la conséquence d’un changement dans leurs écosystèmes naturels (changement climatique, déforestation, urbanisation…).
Une contamination par des animaux vendus sur le marché de Wuhan ayant été fortement suspectée dès le début de l’épidémie, ce marché a été fermé rapidement sans que des prélèvements aient été effectués sur les animaux vivants vendus alors que l’on connaissait le rôle important joué par ces marchés dans l’épidémie du Sras.
Les premières mesures chinoises
Au début de l’apparition de la maladie en Chine, les symptômes observés, rappelant ceux d’une affection grippale avec de la toux et une hyperthermie, n’étaient pas identiques à ceux du Sras et les premiers cas de mortalité rapportés concernaient des personnes âgées ou atteintes d’autres affections sévères. Ceci peut expliquer les premiers propos rassurants de l’OMS alors que les autorités chinoises mettaient en place des mesures de biosécurité particulièrement exceptionnelles (56 millions de personnes confinées dans la province de Hubei, interdiction du commerce d’animaux sauvages, interdiction de voyager, fermeture de la Cité interdite, de la Grande Muraille et des parcs Disney, importantes restrictions de circulation, construction d’hôpitaux dédiés en quelques jours, prolongement du congé du nouvel an chinois, etc.). Ce n’est que le 11 mars 2020 que l’OMS a déclaré qu’il s’agissait d’une pandémie (il s’agit de la première pandémie causée par un coronavirus) [41].
Cependant le virus a pu se propager pendant plus de deux mois en Chine, voire dans d’autres pays, avant la mise en place des mesures de biosécurité drastiques décidées après le 23 janvier 2020, de nombreux déplacements de personnes ayant pu se faire, en particulier avec les préparatifs du nouvel an chinois.
Conclusion
Selon une étude chinoise récente, qui reste à confirmer, portant sur 103 échantillons de virus Sars-CoV-2, il existerait deux souches circulantes, L et S. La souche S (30 % des échantillons) serait plus ancienne que la souche L (70 % des échantillons), celle-ci pouvant être plus agressive et se propager rapidement [42].
Il y aura toujours des maladies émergentes. Une étude effectuée sur 335 maladies ayant émergé entre 1940 et 2008 a permis de noter que 60 % d’entre elles étaient des zoonoses, c’est-à-dire des maladies pouvant infecter l’Homme et l’animal, et parmi celles-ci, 72 % seraient dues à la faune sauvage [43]. C’est pourquoi le concept « une seule santé » est mis en avant par de nombreuses organisations internationales reconnaissant les liens entre la santé humaine, la santé animale et l’environnement [44].
Lors d’une zoonose émergente, il importe aussi de connaître rapidement l’espèce animale qui en est à l’origine ainsi que les autres espèces pouvant jouer le rôle de réservoirs potentiels ou d’hôtes intermédiaires en collectant le plus rapidement possible des prélèvements appropriés avant de pouvoir mettre en place rapidement des mesures efficaces de biosécurité.
La mise en évidence chez les chiroptères de coronavirus pouvant provoquer deux épidémies comme le Sras à partir de 2002 puis le Covid-19 en 2019-2020 doit nous amener à reconsidérer nos stratégies de prévention de ces endémies en évitant le risque de contamination par une cohabitation trop étroite avec les chauves-souris, que ce soit du fait d’une modification de leur écosystème ou de leur consommation.
Il faut aussi éviter les possibilités de transfert de certains virus émergents vers l’Homme par le mélange de plusieurs espèces d’animaux sauvages ou domestiques vendus vivants ou non sur les marchés asiatiques, véritables chaudrons réservoirs de virus et centres d’amplification pour les infections émergentes. Il faut espérer que l’interdiction des marchés d’animaux vivants et plus particulièrement d’animaux sauvages sera maintenue avec rigueur en Chine. Cela impliquera une importante modification des habitudes alimentaires dans plusieurs régions chinoises.
Enfin, les répercussions médicales, économiques et médiatiques de la pandémie due à la maladie Covid-19 démontrent l’intérêt à accorder à l’étude des coronavirus chez les chiroptères mais aussi l’intérêt à protéger ces espèces dont certaines sont insectivores et fort utiles dans la lutte contre les moustiques. L’important est de maintenir un écosystème favorable à tous, animaux sauvages ou domestiques et Homme.
1 | Rodhain F, « Chauves-souris et virus : des relations complexes », Bull. Soc. Pathos. Exot. 2015, 108:272-89.
2 | Calisher CH et al., “Bats : Important reservoir hosts of emerging viruses”, Clin. Microbien. Rev., 2006, 19:531-45.
3 | Middleton DJ et al., “Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus)”, J. Comp. Pathos., 2007, 136:266–72.
4 | Sétien AA et al., “Experimental rabies infection and oral vaccination in vampire bats (Desmodus rotundus)”, Vaccine, 1998, 16:1122-6.
5 | Baker M. et al.,“Antiviral immune responses of bats : A review”, Zoonoses Public Health, 2013, 60:104–16.
6 | Zhang G et al., “Comparative analysis of bat genomes provides insight into the evolution of flight and immunity”, Science, 2013, 339:456–60.
7 | O’Shea TJ et al., “Bat flight and zoonotic viruses”, Emerg. Infect. Dis., 2014, 20:741-7.
8 | Xie J et al., “Dampened STING-Dependent Interferon Activation in Bats”, Cell Host & Microbe, 2018, 23:297-301.
9 | Woo PCY et al., “Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus”, J. Virology, 2012, 3995-4008.
10 | Cheng VC et al., “Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection”, Clin. Microbiol. Rev., 2007, 20:660-94.
11 | Sun RQ et al., “Outbreak of porcine epidemic diarrhea in suckling piglets, China”, Emerg. Infect. Dis., 2012:18:161-3.
12 | Gong LA et al., “New Bat-HKU2–like Coronavirus in Swine, China”, Emerg. Infect. Dis., 2017, 23:167-9. .
13 | OMS, “Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003 (Based on data as of the 31 December 2003)”. Sur www.who.int
14 | OMS, “MERS situation update”, novembre 2019. Sur www.emro.who.int.
15 | Hu B et al., “Discovery of a rich pool of bat SARS-related coronaviruses provides new insights into the origin of SARS”, PLoS Pathogen, 2017, 13.
16 | Fan Y et al., “Bat Coronaviruses in China”, Viruses, 2019, 11::210.
17 | Hu B et al., “Discovery of a rich pool of bat SARS-related coronaviruses provides new insights into the origin of SARS”, PLoS Pathogen, 2017, 13.
18 | Menachery VD et al., “A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence”, Nature Medicine, 2015, 21:1508-13.
19 | Fan Y et al., “Bat Coronaviruses in China”, Viruses, 2019,11:210.
20 | Zhou P et al., “A pneumonia outbreak associated with a new coronavirus of probable bat origin”, Nature, 2020, 579:270-273.
21 | Zhengli Shi Z, Hu Z, “A review of studies on animal reservoirs of the SARS coronavirus”, Virus Research, 2008, 133 : 74-87.
22 | Guan Y et al., “Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China”, Science, 2003, 302:276-8.
23 | Kan B et al., “Molecular evolution analysis and geographic investigation of severe acute respiratory syndrome coronavirus-like virus in palm civets at an animal market and on farms”, J. Virol., 2005, 79:11892-900.
24 | Wang M et al.,“Surveillance on severe acute respiratory syndrome associated coronavirus in animals at a live animal market of Guangzhou in 2004” [Epub ahead of print, in Chinese : Zhonghua Liu Xing Bing Xue Za Zhi, 2005, 26:84-7].
25 | Martina BE et al., “Virology : SARS virus infection of cats and ferrets”, Nature, 2003, 425:915.
26 | Glass WG et al., “Mechanisms of host defense following severe acute respiratory syndrome-coronavirus (SARS- CoV) pulmonary infection of mice”, J. Immunol., 2004, 173:4030-9.
27 | Roberts A et al., “Aged BALB/c mice as a model for increased severity of severe acute respiratory syndrome in elderly humans”, Virol., 2005, 79:5833-8.
28 | Subbarao K et al., “Prior infection and passive transfer of neutralizing antibody prevent replication of severe acute respiratory syndrome coronavirus in the respiratory tract of mice”, J. Virol., 2004, 78:3572-7.
29 | Wentworth DE et al., “Mice susceptible to SARS coronavirus”, Emerg. Infect. Dis., 2004, 10:1293-6.
30 | Liang L et al., “Pathology of guinea pigs experimentally infected with a novel reovirus and coronavirus isolated from SARS patients”, DNA Cell. Biol., 2005, 24:485-490.
31 | Le site de l’International Union for Conservation of Nature (IUCN) : https://www.iucnredlist.org/
32 | Pro-MED-Mail post, “Covid-19 update (37) : China (Hong Kong), animals, dog, preliminary serology negative”, ProMED, 12 March 2020. Sur https://promedmail.org/
33 | Song HD et al., “Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human”, Proc. Nat. Acad. Sci. USA, 2005,102:2430-5.
34 | Wang M et al., “SARS-CoV infection in a restaurant from palm civet”, Emerg. Infect. Dis., 2005, 11:1860-5.
35 | Roberts A et al., “Severe acute respiratory syndrome coronavirus infection of golden Syrian hamsters”, J. Virol., 2005, 79:503-11.
36 | Poon LL et al., “Identification of a novel coronavirus in bats”, J. Virol., 2005, 79:2001-9.
37 | Tu C et al., “Antibodies to SARS coronavirus in civets”, Emerg. Infect. Dis., 2004, 10:2244-8.
38 | Graham RL et al., “A decade after SARS : strategies for controlling emerging coronaviruses”, Nat. Rev. Microbiol., 2013, 11 : 836-48.
39 | Yu H et al., “Effect of closure of live poultry markets on poultry-to-person transmission influenza A H7N9 virus : an ecological study”, Lancet, October 31, 2013.
40 | Wang N et al., “Serological Evidence of Bat SARS-Related Coronavirus Infection in Humans, China”, Virologica Sinica, 2018, 33:104-7.
41 | WHO Director-General’s opening remarks at the media briefing on COVID-19 – 11 March 2020. Sur www.who.int
42 | Tang X et al., “On the origin and continuing evolution of SARS-CoV-2”, National Science Review, nwaa036,
43 | Jones KE et al., “Global trends in emerging infectious diseases”, Nature, 2008, 451:990-3.
44 | OMS, « L’approche multisectorielle de l’OMS “Un monde, une santé” », septembre 2017. Sur www.who.int
Partager cet article




L' auteur

Jeanne Brugère-Picoux
Jeanne Brugère-Picoux est professeure honoraire de l’École nationale vétérinaire d’Alfort, agrégée de pathologie (…)
Plus d'informationsCovid-19

Science, expertise et décision à l’épreuve de la pandémie de Covid-19
Le 15 juillet 2020

Covid-19, hydroxychloroquine et traitement médiatique
Le 9 janvier 2023
Mauvaises conduites et Covid-19
Le 11 octobre 2022
Restaurer l’intégrité scientifique après la crise Covid-19
Le 4 octobre 2022![[16 juin 2022 - Paris] Covid-19 : Deux ans d'épidémie, qu'avons-nous appris ?](local/cache-gd2/0f/33c69087dceb597afeb0e562895a7f.jpg?1675291840)
Communiqués de l'AFIS








